Neúplná dominancia je špeciálny typinterakcia génových alel, v ktorej slabší recesívny znak nemôže byť úplne potlačený dominantným. V súlade so vzormi objavenými G. Mendelom dominantná vlastnosť úplne potláča prejav recesizmu. Výskumník študoval výrazné kontrastné znaky u rastlín s prejavom dominantných alebo recesívnych alel. V niektorých prípadoch sa Mendel stretol s nenaplnením tohto vzoru, ale neuviedol pre to vysvetlenie.
Nová forma dedičstva
Niekedy v dôsledku kríženia potomkovzdedili intermediárne znaky, ktoré rodičovský gén neposkytol v homozygotnej forme. Neúplná dominancia bola v pojmovom aparáte genetiky až začiatkom 20. storočia, keď boli znovuobjavené Mendelove zákony. Zároveň mnohí prírodovedci vykonávali genetické experimenty s rastlinnými a živočíšnymi predmetmi (paradajky, strukoviny, škrečky, myši, ovocné mušky).
Po cytologickom potvrdení Mendelových zákonov Walterom Settonom v roku 1902 sa princípy prenosu a interakcie znakov začali vysvetľovať z hľadiska správania sa chromozómov v bunke.

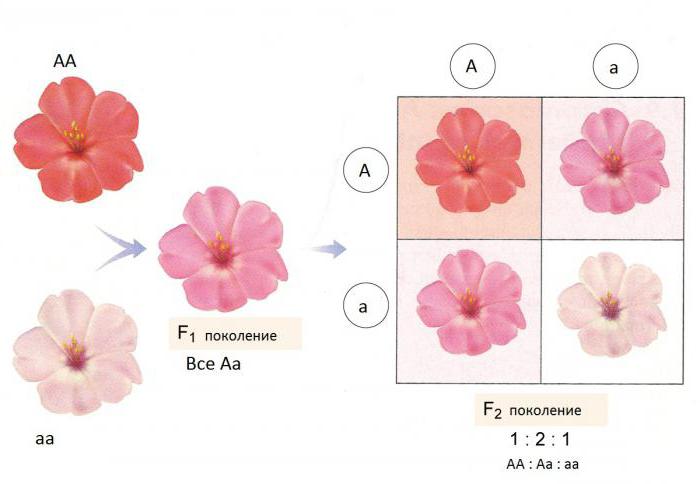
V tom istom roku 1902 opísal prípad Cermak Correnskeď po krížení rastlín s bielymi a červenými korunami malo potomstvo ružové kvety - neúplná dominancia. Tento prejav u hybridov (genotyp Aa) znak, ktorý je stredný vo vzťahu k homozygotnému dominantnému (AA) a recesívne (aa) fenotypy. Podobný účinok bol opísaný pre mnoho druhov kvitnúcich rastlín: snapdragon, hyacint, nočná krása, jahody.
Neúplná dominancia je dôvodom zmeny v práci enzýmov?
Mechanizmus vzhľadu tretieho variantu funkciemožno vysvetliť z hľadiska aktivity enzýmov, ktoré sú svojou povahou proteíny, a gény určujú štruktúru proteínu. V rastline s homozygotným dominantným genotypom (AA) bude dostatok enzýmov a množstvo pigmentu bude správne na intenzívne zafarbenie bunkovej šťavy.
U homozygotov s recesívnymi alelami génu (aa) syntéza pigmentu je narušená, koruna zostáva nenatretá. V prípade intermediárneho heterozygotného genotypu (Aa), dominantný gén stále poskytuje časť enzýmu zodpovedného za pigmentáciu, ale nestačí na jasnú nasýtenú farbu. Ukazuje sa farba "polovičná".
Vlastnosti zdedené stredným typom
Takáto neúplná dedičnosť je dobre sledovaná na znakoch s variabilným prejavom:
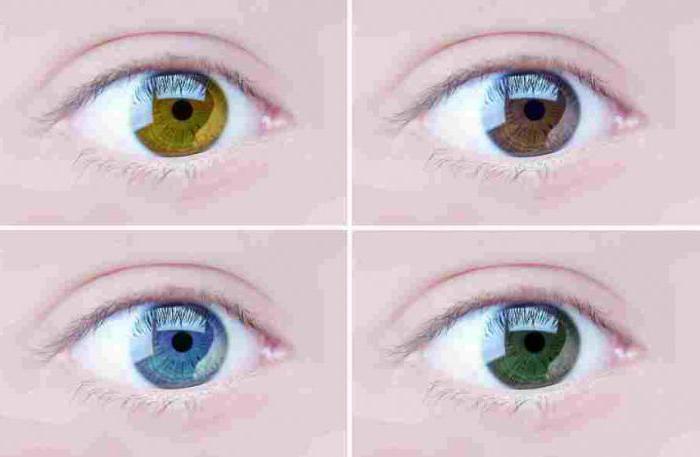
- Intenzita farby. W.Batson krížením čiernych a bielych andalúzskych kurčiat dostal potomkov so strieborným perím. Tento mechanizmus je prítomný aj pri určovaní farby dúhovky ľudských očí.
- Závažnosť vlastnosti. Štruktúra ľudských vlasov je tiež určená neúplným dedením vlastnosti. genotyp AA dáva kučeravé vlasy, aa Sú rovné a ľudia s oboma alelami majú vlnité vlasy.
- Merateľné ukazovatele. Dĺžka pšeničného klasu sa dedí podľa princípu neúplnej dominancie.
V generácii F2 sa počet fenotypov zhoduje s počtomgenotypov, čo charakterizuje neúplnú dominanciu. Analytické kríženia nie sú potrebné na určenie hybridov, pretože sú navonok odlišné od dominantnej čistej línie.
Rozdelenie znakov pri krížení
Úplná a neúplná dominancia ako genetickák interakcii dochádza v súlade s aritmetikou zákonov G. Mendela. V prvom prípade sa pomer fenotypov v F2 (3: 1) nezhoduje s pomerom genotypov potomstva (1: 2: 1), pretože fenotypovo ide o kombinácie alel AA a Aa objavia rovnakým spôsobom. Potom neúplná dominancia je v F2 zhodou podielu rôznych genotypov a fenotypov (1: 2: 1).
U jahôd sa farba dedí rok podľa princípu neúplnej dominancie. Ak skrížite rastlinu s červenými plodmi (AA) a rastlina s bielymi bobuľami - genotyp aa, potom v prvej generácii všetky výsledné rastliny dávajú plody s ružovou farbou (Aa).

Po skrížení hybridov z F1 v druhej generácii F2 dostaneme pomer potomstva, ktorý sa zhoduje s pomerom genotypov: 1AA + 2Aa + 1aa... 25% rastlín z druhej generácie dá červené a nesfarbené plody, 50% rastlín - ružové.
Podobný obraz budeme pozorovať v dvoch generáciách pri krížení čistých línií kvetov nočnej krásky s korunami fialových a bielych kvetov.
Vlastnosti dedičnosti v prípade letality génov
V niektorých prípadoch pomer fenotypovu potomkov je ťažké určiť, ako gény interagujú. V druhej generácii sa rozdelenie s neúplnou dominanciou líši od očakávaného 1: 2: 1 a od 3: 1 - s úplnou dominanciou. Stáva sa to vtedy, keď dominantný alebo recesívny znak dáva v homozygotnom stave fenotyp, ktorý je nezlučiteľný so životom (smrteľné gény).
U sivo sfarbených karakulských oviec umierajú novonarodené jahňatá homozygotné pre dominantnú farebnú alelu v dôsledku skutočnosti, že tento genotyp spôsobuje abnormality vo vývoji žalúdka.
Osoba má príklad letality dominantnej formygén – brachydaktýlia (krátkoprstý). Znak sa zisťuje v prípade heterozygotného genotypu, zatiaľ čo dominantní homozygoti umierajú v skorých štádiách vnútromaternicového vývoja.

Smrteľné môžu byť aj recesívne alely génov.Kosáčikovitá anémia vedie v prípade výskytu dvoch recesívnych alel v genotype k zmene tvaru erytrocytov. Krvné bunky nedokážu účinne pripojiť kyslík a 95 % detí s touto anomáliou zomiera na hladovanie kyslíkom. U heterozygotov zmenená forma erytrocytov neovplyvňuje životaschopnosť do takej miery.
Štiepenie znakov v prítomnosti smrteľných génov
V prvej generácii pri krížení AA x aa letalita sa neprejaví, pretože všetci potomkovia budú mať genotyp Aa. Tu sú príklady rozdelenia znakov v druhej generácii pre prípady so smrteľnými génmi:
Možnosť kríženia Aa x Aa | Úplná nadvláda | Neúplná dominancia |
Dominantná letálna alela | F2: 2 Aa, 1aa Podľa genotypu - 2:1 Podľa fenotypu - 2:1 | F2: 2 Aa, 1aa Podľa genotypu - 2:1 Podľa fenotypu - 2:1 |
Smrteľná alela recesívna | F2: 1AA, 2Aa Podľa genotypu - 1:2 Podľa fenotypu - bez štiepenia | F2: 1AA, 2Aa Podľa genotypu - 1:2 Podľa fenotypu - 1: 2 |
Je dôležité pochopiť, že obe alely pôsobia v prípade neúplnej dominancie a efekt čiastočného potlačenia znaku je výsledkom interakcie génových produktov.









