筋肉の収縮は、以下から構成される複雑なプロセスです。いくつかの段階から。ここでの主成分は、ミオシン、アクチン、トロポニン、トロポミオシン、アクトミオシン、そしてカルシウムイオンと筋肉にエネルギーを供給する化合物です。筋肉収縮の種類とメカニズムを考慮してください。それらがどの段階で構成され、循環プロセスに何が必要かを調べてみましょう。

筋
筋肉は、同じ筋肉収縮メカニズムを持つグループに結合されます。同じ基準で、それらは3つのタイプに分けられます。
- 体の横紋筋;
- 心房および心室の横紋筋;
- 臓器、血管、皮膚の平滑筋。
横紋筋はに含まれていますそれらに加えて、腱、靭帯、骨が含まれているので、その一部である筋骨格系。筋収縮のメカニズムが実装されると、次のタスクと機能が実行されます。
- 体が動く;
- 体の部分は互いに相対的に動きます。
- 体は宇宙で支えられています。
- 熱が発生します。
- 皮質は、受容筋野からの求心によって活性化されます。
それは平滑筋で構成されています:
- 気管支樹、肺および消化管を含む内臓の運動装置;
- リンパ系および循環器系;
- 泌尿生殖器のシステム。
生理学的特性
すべての脊椎動物と同様に、人体は骨格筋繊維の最も重要な3つの特性を区別します。
- 収縮性-興奮時の収縮と緊張の変化;
- 導電率-ファイバー全体の電位の動き。
- 興奮性-膜電位とイオン透過性を変化させることによる刺激への応答。
筋肉が興奮し、から収縮し始めます中心から来る神経インパルス。しかし、人工的な条件では、電気刺激が使用されます。次に、筋肉を直接刺激する(直接刺激)か、筋肉を神経支配する神経を介して刺激する(間接刺激)ことができます。
略語の種類
筋肉収縮のメカニズムは化学エネルギーの機械的仕事への変換。このプロセスは、カエルを使った実験で測定できます。腓腹筋に小さな体重がかかり、軽い電気インパルスで刺激されます。筋肉が短くなる収縮は等張性と呼ばれます。等尺性収縮では、短縮は発生しません。腱は、筋力の発達を短縮することを可能にしません。筋肉収縮の別の補助的メカニズムは、筋肉が最小限の方法で短縮され、強度が最大に発達するときの激しい負荷の状態を前提としています。
骨格筋の構造と神経支配
横紋筋には結合組織に見られ、腱に付着している多くの繊維。いくつかの筋肉では、繊維は長軸に平行ですが、他の筋肉では、それらは斜めの外観を持ち、中央の腱索と羽状のタイプに付着しています。

ファイバーの主な特徴は細いフィラメントの塊の筋形質-筋原線維。それらは、互いに交互になっている明るい領域と暗い領域を含み、隣接するクロスストライプファイバーは同じレベルにあります-断面上にあります。これにより、筋線維全体に沿って横縞が得られます。
サルコメアは暗闇と2つの複合体ですライトディスクで、Z字型の線で区切られています。サルコメアは筋肉の収縮装置です。収縮性筋線維は次のもので構成されていることがわかります。
- 収縮装置(筋原線維システム);
- ミトコンドリア、ゴルジ複合体、および弱い小胞体を備えた栄養装置。
- 膜装置;
- 支援装置;
- 神経装置。
筋線維は、独自の構造と機能を持つ5つの部分に分かれており、筋肉組織の不可欠な部分です。
神経支配
横紋筋におけるこのプロセス線維は、神経線維、すなわち脊髄と脳幹の運動ニューロンの軸索を介して実現されます。 1つの運動ニューロンがいくつかの筋線維を神経支配します。運動ニューロンと神経支配された筋線維との複合体は、神経運動(NME)または運動単位(MU)と呼ばれます。 1つの運動ニューロンが神経支配する繊維の平均数は筋肉MUのサイズを特徴づけ、その逆数は神経支配の密度と呼ばれます。後者は、動きが小さく「薄い」筋肉(目、指、舌)で大きくなります。それどころか、その小さな値は、「荒い」動きのある筋肉(たとえば、体幹)にあります。
神経支配は単一または複数である可能性があります。最初のケースでは、それはコンパクトなモーターエンディングによって実現されます。これは通常、大きな運動ニューロンに当てはまります。筋線維(この場合は物理的または高速と呼ばれます)はAP(活動電位)を生成し、AP(活動電位)がそれらに分配されます。
たとえば、複数の神経支配が発生します。外眼筋で。膜には電気的に興奮するナトリウムチャネルがないため、ここでは活動電位は生成されません。それらでは、脱分極はシナプス終末から線維全体に広がります。これは、筋肉の収縮メカニズムを活性化するために必要です。ここでのプロセスは、最初の場合ほど速くはありません。したがって、それは遅いと呼ばれます。
筋原線維の構造
今日の筋線維の研究は、X線回折分析、電子顕微鏡、および組織化学的方法に基づいて行われています。
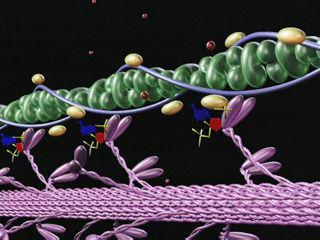
各筋原線維において、直径はこれは1μmで、約2500のプロトフィブリル、つまり細長い重合タンパク質分子(アクチンとミオシン)が含まれています。アクチンプロトフィブリルはミオシンのものより2倍薄いです。安静時、これらの筋肉は、先端のあるアクチンフィラメントがミオシンプロトフィブリルの間の空間に浸透するように配置されています。
ディスクAの細い光の縞模様には、アクチンフィラメントが含まれていません。そして、Zメンブレンはそれらを一緒に保持します。
ミオシンフィラメントに横方向の突起があります長さは最大20nmで、その頭部には約150個のミオシン分子があります。それらは生体極性で出発し、各頭部はミオシンとアクチンフィラメントを接続します。ミオシンフィラメントにアクチン中心の努力があるとき、アクチンフィラメントはサルコメアの中心に近づきます。最後に、ミオシンフィラメントはZ線に到達し、サルコメア全体を占め、アクチンフィラメントはそれらの間にあります。この場合、ディスクIの長さが短くなり、最終的には完全に消え、それに伴って線Zが太くなります。
それで、滑り糸の理論によれば、筋線維の長さの減少が説明されます。 「歯車」と呼ばれるこの理論は、20世紀半ばにハクスリーとハンソンによって開発されました。

筋線維収縮のメカニズム
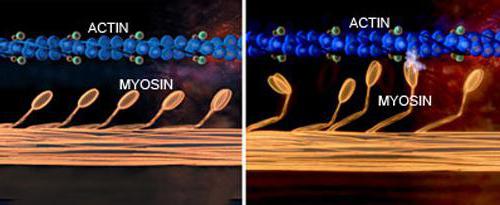
理論上の要点は、スレッドがないということです(ミオシンとアクチン)が短縮されます。筋肉が伸ばされても、それらの長さは変わりません。しかし、細い糸の束が滑って太い糸の間に出て、重なりの度合いが減り、収縮が起こります。
筋収縮の分子メカニズムアクチンフィラメントをスライドさせることにより、以下のようになります。ミオシンヘッドはプロトフィブリルとアクチンを接続します。それらが傾くと、スリップが発生し、アクチンフィラメントがサルコメアの中心に向かって移動します。フィラメントの両側にミオシン分子が双極性に組織化されているため、アクチンフィラメントをさまざまな方向にスライドさせるための条件が作成されます。
筋肉が弛緩すると、ミオシンの頭がアクチンフィラメントから離れます。簡単にスライドできるため、リラックスした筋肉はストレッチに抵抗しません。したがって、それらは受動的に長くなります。
削減段階
筋収縮のメカニズムは、以下の段階に簡単に細分できます。
- 活動電位がシナプスの運動ニューロンに由来する場合、筋線維が刺激されます。
- 活動電位は筋線維の膜で生成され、筋原線維に広がります。
- 電気機械結合が実行されます。これは、電気PDを機械的スライドに変換することです。カルシウムイオンは必然的にこれに関与します。
カルシウムイオン

繊維活性化プロセスをよりよく理解するためにカルシウムイオンを含むアクチンフィラメントの構造を考えると便利です。その長さは約1μm、その厚さは5〜7nmです。これは、アクチンモノマーに似た一対のねじれたストランドです。ここには約40nmごとに球状のトロポニン分子があり、鎖の間にトロポミオシン分子があります。
カルシウムイオンが存在しない場合、それは筋原線維は弛緩し、長いトロポミオシン分子はアクチン鎖とミオシンブリッジの付着をブロックします。しかし、カルシウムイオンが活性化されると、トロポミオシン分子はより深く下がって、領域が開きます。
次に、ミオシンブリッジが接続しますアクチンフィラメント、ATPが分解され、筋力が発達します。これは、トロポニンに対するカルシウムの作用によって可能になります。この場合、後者の分子が変形し、それによってトロポミオシンが押し出されます。
筋肉が弛緩すると、湿重量1グラムあたり1μmol以上のカルシウムが含まれます。カルシウム塩は分離され、特別な保管施設に保管されます。そうでなければ、筋肉は常に収縮します。
カルシウムの貯蔵は以下の通りです。繊維の内側の筋細胞の膜のさまざまな部分に、細胞の外側の環境との接続が行われるチューブがあります。横管系です。そしてそれに垂直に縦方向のシステムがあり、その端には横方向のシステムの膜のすぐ近くに位置する気泡(末端槽)があります。一緒に彼らはトライアドを作ります。カルシウムが蓄えられるのは泡の中にあります。
これは、PDがセルに広がる方法です。電気機械結合が発生します。興奮は繊維に浸透し、縦方向のシステムに入り、カルシウムを放出します。したがって、筋線維の収縮のメカニズムが実行されます。
ATPによる3つのプロセス
カルシウムイオンの存在下での両方の鎖の相互作用において、ATPは重要な役割を果たします。骨格筋の筋収縮のメカニズムが実装されている場合、ATPエネルギーは次の目的で使用されます。
- 一定濃度のイオンを維持するナトリウムおよびカリウムポンプの操作。
- 膜の反対側にあるこれらの物質;
- 筋原線維を短縮するスライディングスレッド;
- リラックスする働きをするカルシウムポンプの働き
ATPは、細胞膜、ミオシンフィラメント、筋小胞体の膜に見られます。酵素はミオシンによって切断され、利用されます。
ATP消費
ミオシンヘッドが相互作用することが知られていますアクチンとATPの切断のための要素が含まれています。後者は、マグネシウムイオンの存在下でアクチンとミオシンによって活性化されます。したがって、酵素の切断は、ミオシンヘッドがアクチンに付着したときに発生します。さらに、横方向のブリッジが多いほど、分割率が高くなります。
ATPメカニズム
運動の終了後、AFT分子反応に関与するミオシンとアクチンを分離するためのエネルギーを提供します。ミオシンヘッドは分離され、ATPはリン酸とADPに分解されます。最後に、新しいATP分子が付着し、サイクルが再開されます。これが分子レベルでの筋肉の収縮と弛緩のメカニズムです。
クロスブリッジの活動は、ATPの加水分解が起こっている間だけ継続します。酵素がブロックされると、ブリッジは再付着しません。
生物の死が始まると、細胞内のATPのレベルが低下し、ブリッジはアクチンフィラメントに安定して付着したままになります。これは死後硬直の段階です。

ATPの再合成
再合成は2つの方法で実装できます。
からの酵素伝達を介してADPのリン酸基のクレアチンリン酸。細胞内のクレアチンリン酸の貯蔵量はATPよりもはるかに多いため、再合成は非常に迅速に実現されます。同時に、ピルビン酸と乳酸の酸化によって再合成がゆっくりと進行します。
再合成するとATPとCPが完全に消失する可能性があります毒に邪魔されます。その後、カルシウムポンプが機能を停止し、その結果、筋肉が不可逆的に収縮します(つまり、拘縮が起こります)。したがって、筋肉の収縮のメカニズムが破壊されます。
プロセス生理学
上記を要約すると、次のことに注意してください。筋線維の収縮は、各サルコメアの筋原線維を短くすることから成ります。ミオシン(太い)とアクチン(細い)のフィラメントは、リラックスした状態で両端で接続されています。しかし、筋肉の収縮のメカニズムが実現されると、それらは互いに向かってスライド運動を開始します。生理学は、ミオシンの影響下で、ATPをADPに変換するために必要なエネルギーが放出されるプロセスを(簡単に)説明します。この場合、ミオシンの活性は、筋小胞体に蓄積するカルシウムイオンの十分な含有量によってのみ実現されます。





